
Laura Zanin1 , Nicola Tomasi1 , Stefano Cesco2 , Zeno Varanini3 and Roberto Pinton1
1 Dipartimento di Scienze AgroAlimentari, Ambientali e Animali, Università degli Studi di Udine, Udine, Italy, 2 Faculty of Science and Technology, Free University of Bozen-Bolzano, Bolzano, Italy, 3 Dipartimento di Biotecnologie, Università di Verona, Verona, Italy
บทคัดย่อ
งานวิจัยจำนวนมากพบว่าสารอาหารธาตุเหล็กพืชที่นำไปใช้ได้เพิ่มขึ้น จากการก่อรูปเป็นโครงสร้างเชิงซ้อนของธาตุโลหะต่าง ๆ ที่จับคู่กับสารฮิวมิค อันมีที่มาจากแหล่งต่างกัน โดยทั่วไป อินทรีย์วัตถุจะแปรสภาพเป็นฮิวมิคในดินตะกอนและสารละลาย และมีบทบาทในการเพิ่มการสะสมธาตุเหล็ก (Fe) ในดินและพืช โดยปริมาณการสะสมนี้ ขึ้นอยู่กับประสิทธิภาพการละลายน้ำและขนาดโมเลกุลของฮิวมิค ซึ่งจะคายลิแกนด์โลหะ (metal ligand) ออกมา ทั้งนี้ฮิวมิคยังเอื้อให้โครงสร้างเชิงซ้อนระหว่างธาตุเหล็ก (Fe) และฮิวมิค อยู่ในสภาพที่พร้อมใช้งานสำหรับกระบวนการดูดซึมธาตุเหล็กของพืช งานวิจัยยังพบด้วยว่าฮิวมิคจะเพิ่มศักยภาพทางกายภาพของกระบวนการดูดซึมแร่ธาตุที่เกี่ยวข้อง โดยธาตุเหล็ก (Fe) จะมีบทบาททั้งในช่วง Transcriptional และ Post-Transcriptional ของการสังเคราะห์ RNA พืช อย่างไรก็ตามการกระจายและการเคลื่อนย้ายธาตุเหล็ก (Fe) ในส่วนต่าง ๆ ของพืช ยังอาจถูกดัดแปลงได้ หากพืชได้รับคีเลต (โครงสร้างเชิงซ้อน) ธาตุเหล็ก-ฮิวมิค ที่ละลายน้ำได้ และมีโครงสร้างแบบเดียวกันกับคีเลตของธาตุอื่น ๆ (ทั้งสังเคราะห์หรือเกิดตามธรรมชาติ) ผลการวิจัยนี้ ตรงกับที่เคยสังเกตในงานวิจัยก่อน ๆ ซึ่งระบุว่า การเติมฮิวมิคเข้าไป สามารถดัดแปลงโครงสร้างรากพืชและยังกำหนดกิจกรรมในชั้นเนื้อเยื่อเซลล์พืช (plant membrane) ที่เกี่ยวข้องกับเส้นทางการลำเลียงสารอาหารของระบบการเผาผลาญพลังงานชั้นเอก (Primary) และชั้นรอง (Secondary) และส่งผลต่อสมดุลฮอร์โมน กับสมดุลปฏิกิริยาออกซิเจน บทบาทของฮิวมิคที่หลากหลายบ่งชี้ว่า โครงสร้างเชิงซ้อนของฮิวมิคและธาตุเหล็ก (Fe) ที่ละลายน้ำได้ สนับสนุนกระบวนการดูดซึมของพืชในแบบที่ซับซ้อน โดยการจัดหาธาตุเหล็กในรูปแบบที่พร้อมใช้งานในไรโซเสฟียร (rhizosphere) และโดยการกระทบต่อโครงสร้างทางกายภาพพืชโดยตรง นอกจากนี้ อาจสามารถพิจารณาได้ว่า การมีธาตุเหล็ก (Fe) และฮิวมิคที่สามารถละลายน้ำได้ จากหลาย ๆ แหล่ง หลาย ๆ ขนาดโมเลกุล ในรูปแบบสารละลายที่ต่างกัน เพื่อใช้เป็นปุ๋ยธาตุเหล็ก (Fe) ถือเป็นการอนุรักษ์สิ่งแวดล้อมในการเกษตรแบบหนึ่ง
คำสำคัญ: โครงสร้างธาตุเหล็กเชิงซ้อน (Fe) คีเลตธาตุเหล็ก (Fe) กรดฟุลวิค, การดูดซึมทางราก กลยุทธ์ที่ 1 กลยุทธ์ที่ 2 สารละลายฮิวมิคดึงน้ำออกได้ (water-extractable humic substances-WEHS)
บทนำ
ดินที่มีฮิวมิคคือดินที่เกิดจากการสลายตัวบางส่วนของหินและการสังเคราะห์อินทรีย์วัตถุขึ้นมาใหม่ โดยเฉพาะอินทรีย์วัตถุจากพืช ดินที่มีฮิวมิคเกิดจากกระบวนการพอลิเมอไรเซชันและพอลิคอนเดนเซชัน (polymerization / polycondensation) ของสารประกอบฟีนอล ที่ได้จากการย่อยสลายของลิกนินแบคทีเรีย ดังนั้น ดินที่มีฮิวมิคจึงมีกลิ่นหอมเองตามธรรมชาติ กระบวนการนี้นำโมเลกุลอินทรีย์หลาย ๆ ชนิด รวมถึงโมเลกุลสายโซ่อะลิฟาติก (คือ สารประกอบไฮโดรคาร์บอน ที่ภายในโมเลกุลมีอะตอมคาร์บอนต่อกันเป็นลูกโซ่) โมเลกุลเปปไทด์ โมเลกุลกรดอะมิโน กรดไขมัน และโมเลกุลน้ำตาล มารวมกันเป็นโมเลกุลขนาดใหญ่ ที่มีน้ำหนักโมเลกุลปานกลาง-มาก (Stevenson, 1994) นอกจากนี้ดินฮิวมิค ยังเกิดจากโมเลกุลฮิวมิคขนาดเล็กเชื่อมโยงเข้าด้วยกันผ่านปฏิกิริยาไฮโดรโฟบิกและพันธะไฮโดรเจนได้อีกด้วย (Piccolo, 2002) โมเลกุลฮิวมิคที่มีมวลโมเลกุลต่างกันสามารถทำพันธะระหว่างกันได้ เป็นโครงข่ายโมเลกุลฮิวมิคขนาดใหญ่ โดยขนาดของโครงข่ายจะขึ้นอยู่กับค่า pH และกำลังของพันธะไอออนิค และแร่ธาตุที่เป็นองค์ประกอบในสารละลาย (Garcia-Mina, 2007; Esfahani et al., 2015)
กระบวนการเหล่านี้บ่งบอกเป็นนัยว่าในดินมีฮิวมิคที่มีขนาดโมเลกุลต่างกัน และมีศักยภาพการละลายต่างกัน โดยจะปรากฏเศษโมเลกุลบางส่วนปรากฏในสารละลายดิน จึงสามารถทำปฏิกิริยากับรากพืชได้โดยตรง (Chen and Schnitzer, 1978; Gerke, 1997) สารละลายที่ฮิวมิคที่ปรากฏในดินเหล่านี้ ถือเป็นสารละลายอินทรีย์วัตถุประเภทหนึ่ง (Dissolved Organic Matter-DOM) (Bolan et al., 2011)
โดยปรกติสามารถใช้สารละลายด่างชนิดต่าง ๆ ดึงแยกฮิวมิคจากดินได้ ซึ่งโมเลกุลสามารถนำมาเข้ากระบวนการแยกสารประกอบออกจากันได้อีกขั้นหนึ่ง เป็นกรดฮิวมิคประเภทต่าง ๆ และกรดฟุลวิก แบ่งตามเกณฑ์ศักยภาพการละลายน้ำ (Stevenson, 1994)
เนื่องจากโครงสร้างโมเลกุลที่มีความหลากหลาย จึงไม่สามารถระบุลักษณะเฉพาะของโครงสร้างโมเลกุลดินที่มีฮิวมิคได้อย่างชัดเจน อย่างไรก็ตามมีการระบุอย่างชัดเจนว่าฟังชันนอลกรุ๊ป ภายในโครงสร้างโมเลกุล มีผลทางตรงและทางอ้อมต่อการเจริญเติบโตของพืชและสารอาหาร (Nebbioso and Piccolo, 2011; Muscolo et al., 2013; García et al., 2016a) ผลทางอ้อมหมายถึงการเปลี่ยนแปลงองค์ประกอบทางเคมีและกายภาพของดินและไรโซสเฟียร์ ขณะที่ผลทางตรงคือกิจกรรมในชั้นเนื้อเยื่อพลาสมาเมมเบรน (Plasma Membrane- PM) และเส้นทางการเผาผลาญพลังงานของพืช (Varanini and Pinton, 2001; Nardi et al., 2002; Zandonadi et al., 2013; Canellas and Olivares, 2014; Rose et al., 2014; Olaetxea et al., 2018)
มีการสันนิฐานเกี่ยวกับการปรากฏฮิวมิคในดิน เนื่องจากฮิวมิคเป็นตัวแทนของวิวัฒนาการขั้นสูงตามธรรมชาติทางชีววิทยาของอินทรีย์วัตถุ ว่าเป็นผลของกระบวนการดึงแยกฮิวมิค โดยใช้กระบวนการของสารละลายด่าง (Lehmann and Kleber, 2015) อย่างไรก็ตามข้อสันนิฐานนี้ยังคงเป็นที่ถกเถียงอยู่ (Gerke, 2018; Olk et al., 2019) ทั้งนี้เป็นที่น่าสังเกตว่ามีงานวิจัยที่เคยเฝ้าสังเกตพบปรากฎการณ์ การดึงโมเลกุลคล้ายฮิวมิคจากดิน ที่เคยผ่านการใช้ของเหลวความเข้มข้นอ่อน ๆ ในการดึงแยกฮิวมิคออกมา (Hayes, 2006) ทั้งนี้ปรากฎการณ์นี้พบได้เฉพาะในสภาพแวดล้อมทางน้ำ (Aquatic Environments) (Alberts and Takács, 2004) เช่นฮิวมิคในพื้นดินที่มีน้ำขัง (peat water) และตะกอนที่พัดมากับน้ำ (soil leachate) (Pinton et al., 1997; Vujinovic et al., 2013)
แม้ว่ายังมีข้อกังขาเกี่ยวกับสถานภาพทางเคมีของฮิวมิค แต่ก็มีการแสดงให้เห็นอย่างแจ่มชัดว่าอินทรียวัตถุจากแหล่งที่มาที่ต่างกันสามารถให้ธาตุเหล็ก (Fe) ในรูปที่พืชสามารถใช้งานได้ โดยเป็นผลของการก่อตัวของโครงสร้างเชิงซ้อนระหว่างธาตุเหล็ก (Fe) กับโมเลกุลฮิวมิค (Chen et al., 2004a; Bocanegra et al., 2006; Kovács et al., 2013; Cieschi et al., 2017) นอกจากนี้ยังมีก่อตัวของโครงสร้างเชิงซ้อนละลายน้ำได้ของธาตุเหล็กและฮิวมิค (Fe-Hs) ซึ่งพืชสามารถนำไปใช้ได้เลยโดยตรง (Pandeya et al., 1998; Pinton et al., 1999) การที่ฮิวมิคสามารถก่อตัวกับธาตุโลหะต่าง ๆ เป็นโครงสร้างเชิงซ้อนได้ และมีผลกระทบต่อกระบวนการดูดซึมอาหารและการเผาผลาญพลังงานของพืชนี้ เป็นเครื่องบ่งชี้บทบาทที่หลากหลายของเศษอินทรีย์วัตถุที่มีต่อสารอาหารธาตุเหล็ก (Fe)
ผลของฮิวมิคต่อธาตุเหล็ก
ฮิวมิค เมื่อรวมกับโมเลกุลสารอาหารโลหะขนาดเล็ก สามารถก่อเป็นโครงสร้างเชิงซ้อนที่มีความเสถียรได้ เนื่องจากภายในโมเลกุลมีโครงสร้างออกซิเจน-ไนโตรเจน-ซัลเฟอร์ ที่มีฟังชันนอลกรุ๊ป (Functional Groups) ซึ่งจะช่วยรักษาสภาพสารอาหารขนาดเล็กในสารละลาย และ หรือ รักษาสภาพสารอาหารขนาดเล็กที่ปรากฏในดินทุกประเภท ณ ค่า pH ธรรมชาติ (Senesi, 1992; Tipping, 2002) ในกรณีของโมเลกุลโลหะธาตุเหล็ก (Fe) กับฮิวมิค กระบวนการก่อตัวจะมีการทำปฏิกิริยากับกลุ่มโมเลกุลธาตุออกซิเจน (กลุ่มกรดคาร์บอกซิลิก และสารประกอบฟีนอล) (Senesi, 1992; Tipping, 2002) ทั้งนี้งานวิจัยเมื่อเร็ว ๆ นี้ พบว่ากรดคาร์บอกซิลิกในสารประกอบแอลิแฟติก (aliphatic domains) มีส่วนในกระบวนการก่อตัวระหว่าง Fe(III) กับฮิวมิค (Fuentes et al., 2013)
การประเมิณความเสถียรของลำดับโครงสร้างเชิงซ้อนที่ก่อตัวระหว่างธาตุโลหะต่าง ๆ และกรดฮิวมิคต่าง ๆ กระทำโดยใช้วิธีพเทนชิโอเมตริกไทเทรชันและ Irving-Williams series (Garcia-Mina et al., 2004) โดยค่าที่พบได้ มีค่าต่ำกว่าที่สังเกตได้ในโครงสร้างเชิงซ้อนระหว่างธาตุเหล็ก (Fe) และตัวทำปฏิกิริยาคีเลตแบบสังเคราะห์ (e.g., EDTA, EDDHA; Lucena, 2003) หรือที่สังเกตได้ในสารประกอบอินทรีย์ที่พบในธรรมชาติ (เช่น กรดอินทรีย์ต่าง ๆ ซิเดอร์โรฟอร์, ไฟโทซิเดโรฟอร์, สารประกอบฟีนอล) (von Wirén et al., 2000; Crowley, 2001; Ryan et al., 2001; Mimmo et al., 2014) ทั้งนี้ความเสถียรและศักยภาพการละลายของโครงสร้างเชิงซ้อนนี้เปลี่ยนแปลงได้ หากค่า pH และอัตราส่วนโมลาร์ (Molar Ratio) ระหว่างสารอาหารขนาดเล็กกับฮิวมิคเปลี่ยนไป (Chen et al., 2004a; Garcia-Mina, 2006).
ในช่วง pH 5 – 9 ความเสถียรของโครงสร้างเชิงซ้อนจะเพิ่มขึ้นหากมีอัตราส่วนโมลาร์ (Molar Ratio) ระหว่างธาตุโลหะและสสารฮิวมิคต่ำ ขณะที่ศักยภาพการละลายจะเพิ่มขึ้นตามค่าความเป็นด่าง และเมื่อมีอัตราส่วนโมลาร์ (Molar Ratio) ระหว่างธาตุเหล็กและฮิวมิคที่ต่ำ ซึ่งอัตราส่วนนี้บอกเป็นนัยว่าพืชที่ปลูกในดินเนื้อปูน ที่มีปริมาณธาตุเหล็ก (Fe) จำกัดจะได้ประโยชน์จากความเสถียรในการก่อตัวของโครงสร้างสารละลายได้เชิงซ้อนอย่าง โครงสร้างธาตุเหล็ก (Fe) – ฮิวมิค (Cieschi and Lucena, 2018) และได้ประโยชน์จากโครงสร้างเชิงซ้อนที่ละลายไม่ได้ แต่มีน้ำหนักโมเลกุลมาก อย่างฮิวมิค (Colombo et al., 2014)
ฮิวมิคมีผลต่อปริมาณที่ปรากฏของธาตุเหล็ก (Fe) โดยน้ำหนักของเศษเสี้ยวต่าง ๆ ของโมเลกุลฮิวมิค จะไปกระทบต่อเสถียรภาพของการกระจายตัวของธาตุเหล็กออกไซด์รูปอสัณฐาน (amorphous Fe oxides; poorly crystalline; crystalline phase) (Schwertmann, 1991) ธาตุเหล็กที่ก่อตัวเป็นรูปอสัณฐานนี้ตกตะกอนร่วมกับฮิวมิคที่ไม่ละลาย (Insoluble Humic Substance) และคงสภาพไว้ในรูปนี้อย่างยาวนาน สะสมไว้ในรูปลิแกนด์ (Ligand) กล่าวคือ สะสมไว้โดยการจับคู่พันธะระหว่างอะตอมฮิวมิคกับธาตุเหล็ก โดยจับเอาธาตุเหล็กเป็นแกนพันธะ เพื่อการเตรียมเคลื่อนย้าย (ligand mobilization) ให้พืชดูดซึม (Colombo et al., 2012, 2014)
ศักยภาพของฮิวมิคต่อการก่อโครงสร้างเชิงซ้อนกับธาตุเหล็กเองก็มีความสำคัญกับการจัดหาสารอาหารฟอสฟอรัสให้กับพืช เนื่องจากฟอสฟอรัสทำพันธะกับฮิวมิคโดยใช้ธาตุเหล็ก (Fe) เป็นตัวกลาง (Gerke, 2010; Urrutia et al., 2013) กระบวนการนี้จะเพิ่มปริมาณฟอสเฟตที่พืชสามารถใช้งานได้ อันที่จริง การก่อโครงสร้างเชิงซ้อนของธาตุเหล็ก (Fe) ในรูปแบบ Ligand mobilization มีส่วนมาจากการกระตุ้นของรากพืชที่ปล่อยสารคัดหลั่งจากราก เพื่อกระตุ้นกิจกรรมของแบคทีเรียในดิน และส่งเสริมการดูดซึมสารอาหารทั้งสองประเภทไปยังราก (Gerke, 1993; Urrutia et al., 2014)
ฮิวมิคมักนำให้เกิดปฏิกิริยา Redox Reactive (Oxidation-Reduction) และสามารถดึงเอาไอออนจากธาตุโลหะอย่าง Fe3+ (Skogerboe and Wilson, 1981; Struyk and Sposito, 2001) การดึงไอออนจากธาตุโลหะนี้เกิดขึ้นอย่างมีนัยยะสำคัญเมื่อค่า pH ต่ำกว่า 4 เพราะเมื่อค่า pH สูงกว่า 4 ปฏิกิริยา Redox Reactive นี้จะโดนจำกัดโดยการก่อตัวของโครงสร้างเชิงซ้อนระหว่าง Fe3+ กับโมเลกุลฮิวมิค ทั้งนี้งานวิจัยได้แสดงให้เห็นว่าฮิวมิคที่ถูกละลายแล้วและที่ยังอยู่ในสถานะของแข็ง สามารถเร่งให้เกิดการตกตะกอนได้ (Nevin and Lovley, 2002; Roden et al., 2010) และยังทำปฏิกิริยาทางชีวภาพในการดึงไอออนจากแร่โลหะเหล็ก Fe(III) ในดิน
บทบาทของฮิวมิคในฐานะของคีเลต (พาหะ) จากธรรมชาติ
ฮิวมิคนอกจากจะชะลอกระบวนการก่อรูปสัญฐานของธาตุเหล็ก (Fe) แล้ว ยังมีส่วนช่วยในการก่อรูปโครงสร้างเชิงซ้อนธาตุเหล็ก (Fe) – ฮิวมิค แบบสารละลายที่พืชสามารถดูดเข้าไปใช้ในราก (Pandeya et al., 1998; Garcia-Mina et al., 2004; Chen et al., 2004b) โครงสร้างเชิงซ้อนนี้ทำหน้าที่เป็นคีเลต (พาหะ) ที่เกิดตามธรรมชาติของธาตุเหล็ก (Fe) ซึ่งทำปฏิกิริยากับกระบวนการดูดซึมของพืช โดยอาศัยเศษฮิวมิคที่สามารถดึงแยกโมเลกุลน้ำได้ (WEHS) purified from a water extract of sphagnum peat มีการพบว่าสามารถก่อรูปโครงสร้างเชิงซ้อนระหว่างธาตุเหล็ก (Fe) กับ เศษฮิวมิคที่สามารถดึงแยกโมเลกุลน้ำได้ (WEHS) โดยธาตุเหล็ก (Fe) อยู่ในรูปโครงสร้างที่ละลายน้ำไม่ดี (Cesco et al., 2000) ทั้งนี้พืชที่มีลักษณะขาดธาตุเหล็ก (Fe) ตามกลุ่ม Strategy I และ Strategy II สามารถนำโครงสร้างเชิงซ้อนธาตุเหล็ก-ฮิวมิคที่ดึงแยกโมเลกุลน้ำได้นี้ ไปใช้ได้ โดยพืชในกลุ่ม Strategy I จะใช้กระบวนการดูดซึมแบบ Fe(III) reduction-based (Pinton et al., 1999) ส่วนพืชในกลุ่ม Strategy II ใช้การแลกเปลี่ยนลิแกนด์ (Ligand) ระหว่างฮิวมิคที่ดึงแยกโมเลกุลน้ำได้ กับไฟโตไซเดอโรฟอร์ (PS) (Cesco et al., 2002) การตรวจวัดค่าการดูดซึม 59Fe จากโครงสร้างเชิงซ้อน 59Fe-WEHS นั้นใช้ค่า pH เปรียบเทียบกับค่า pH ที่พบในดินเนื้อปูน (calcareous soils) (Cesco et al., 2002; Tomasi et al., 2013) ทั้งนี้ การดูดซึมโดยรากของพืชในกลุ่ม Strategy I ก็ใช้วิธีการตรวจวัดแบบเดียวกัน (Tomasi et al., 2013; Zamboni et al., 2016) เมื่อพืชที่ขาดธาตุเหล็ก (Fe) ได้รับโครงสร้างเชิงซ้อนธาตุเหล็ก-ฮิวมิคที่ละลายน้ำได้ (Fe-WEHS) ก็มีการปรับฟื้นตัวที่เทียบเคียงได้กับการกระตุ้น of the acidification capacity of roots, ซึ่งเป็นองค์ประกอบหนึ่งในกนการตอบสนองต่อภาวะขาดธาตุเหล็ก (Fe) ในพืชกลุ่ม Strategy-I (Pinton et al., 1999; Tomasi et al., 2013)
พืชมีการสะสมธาตุเหล็กจากคีเลตโครงสร้างเชิงซ้อนธาตุเหล็ก-ฮิวมิคที่ดึงแยกน้ำออกได้ (59Fe-WEHS) มากกว่าการสะสม คีเลตโครงสร้างเชิงซ้อนอื่น ๆ เช่นธาตุเหล็ก-ไซเตรต (59Fe-Citrate) หรือธาตุเหล็ก-ไฟโตไซเดอโรฟอร์ (59Fe- PS) (Tomasi et al., 2013; Zamboni et al., 2016) นอกจากนี้ ยังพบอัตราการเคลื่อนย้ายคีเลตธาตุเหล็ก (Fe) ไปยังใบที่สูงกว่าคีเลตธาตุอื่น ๆ ในพืชที่ขาดธาตุเหล็ก (Fe) ในกลุ่ม Strategy-I โดยพืชในกลุ่มนี้ทั้งหมด ได้รับโครงสร้างเชิงซ้อนธาตุเหล็ก-ฮิวมิคที่ดึงแยกน้ำออกได้ (59Fe-WEHS) (Tomasi et al., 2009; Zanin et al., 2015) การเคลื่อนย้ายคีเลตธาตุเหล็กที่สูงกว่าคีเลตธาตุอื่น ๆ นี้ ตามมาด้วยการเพิ่มขึ้นของปริมาณธาตุเหล็กในท่อลำเลียงไซเล็ม (Tomasi et al., 2009) ยกตัวอย่าง เช่น การทดลองในพันธุ์พืชแตงกวาที่ได้รับคีเลตธาตุเหล็ก-ฮิวมิคที่ดึงแยกน้ำออกได้ (59Fe-WEHS) โดยพบว่าธาตุเหล็ก (Fe) ในต้นพืช เคลื่อนย้ายไปยังใบและต่อไปยังเซลล์จัดเก็บระหว่างท่อลำเลียง (Interveinal Cells) (Zanin et al., 2015) Bocanegra et al. (2006) ทำการทดลอง โดยสังเกตว่ามีการเคลื่อนย้ายธาตุเหล็ก (Fe) ที่รวดเร็วจากรากไปยังใบของพืชที่ได้รับเศษโมเลกุลฮิวมิคน้ำหนักโมเลกุลน้อย ผลทดลองนี้ชี้ให้เห็นว่าฮิวมิคมีผลต่อการได้รับสารอาหารธาตุเหล็ก (Fe) ของพืช ผ่านการเพิ่มปริมาณธาตุโลหะในดินและในไรโซสเฟียร์ นอกจากนี้ฮิวมิคยังมีผลต่อกระบวนการดูดซึมและการเคลื่อนย้ายสารอาหารภายในพืช
มีงานวิจัยที่แสดงว่า ฮิวมิคและคีเลตโครงสร้างเชิงซ้อนธาตุเหล็ก-ฮิวมิค (Fe-HS) มีผลต่อยีนที่เกี่ยวข้องกับกระบวนการดูดซึมธาตุเหล็ก (Fe) ของพืช โดยพืชดูดซึมโครงสร้างเชิงซ้อนธาตุเหล็ก-ฮิวมิคที่ดึงแยกน้ำออกได้ (Fe-WEHS) เช่น ในต้นมะเขือเทศ การให้ปุ๋ยโครงสร้างเชิงซ้อนฮิวมิค-ธาตุเหล็ก (HS-Fe Complexes) ส่งเสริมการดูดคีเลตธาตุเหล็ก Fe(III) ขึ้นไปจากราก และมีผลต่อ reductase (LeFRO1) and Fe transporter genes, LeIRT1 and LeIRT2 (Tomasi et al., 2013) การเพิ่มขึ้นของ transcript abundance นั้นรวดเร็วกว่า และถึงระดับที่สูงกว่าเมื่อให้ปุ๋ยโครงสร้างเชิงซ้อนธาตุเหล็ก-ไซเตรต (Fe-Citrate) หรือปุ๋ยธาตุเหล็ก -ไฟโตไซเดอโรฟอร์ (Fe-PS) งานวิจัยของ Aguirre et al. (2009) ชี้ให้เห็นว่าการใส่ฮิวมิคที่ถูกทำให้บริสุทธิ์จาก leonardite induced a transient up-regulation of genes ที่ทำหน้าที่เกี่ยวข้องกับกระบวนการดูดซึมของพืชในกลุ่ม Strategy I กล่าวคือ ยีน CsHA2, CsFRO1 และ CsIRT1 ในรากต้นแตงกวา ผลลัพธ์เหล่านี้ล้วนเชื่อมโยงกับการเพิ่มขึ้นของ Fe(III) chelate-reductase activity ในราก Billard et al. (2013) ได้ทำการวิจัยเศษฮิวมิคที่แยกจาก black peat และค้นพบว่าเศษฮิวมิคมีผลต่อยีน IRT1 ซึ่งเป็นยีนที่เกี่ยวข้องกับการดูดซึมขึ้นลำต้นไปยังใบของผักกาดก้านขาว ผลงานวิจัยนี้มีความสัมพันธ์กันอย่างมีนัยะสำคัญต่อปริมาณความเข้มข้นของธาตุเหล็ก (Fe) ในใบ
เป็นที่น่าสนใจว่ายีนที่เกี่ยวข้องกับการดูดซึมธาตุเหล็ก (Fe) ในใบ (CsFRO1, CsIRT1, CsNRAMP) ) ควบคุมการดูดสารอาหารขึ้นตามปริมาณปุ๋ยโครงสร้างเชิงซ้อนธาตุเหล็ก-ฮิวมิคที่ดึงน้ำออกได้ (Fe- WEHS) ที่ให้กับพืชตระกูลแตงกวาที่มีภาวะขาดธาตุเหล็ก (Fe) (Zanin et al., 2015) The localization of CsFRO1, CsIRT1 transcripts was evident next to the midveins, while CsNRAMP expression was detected in the overall mesophyll region, supporting a role of this later gene ในการกระจายธาตุเหล็ก (Fe) ไปตลอดเนื้อเยื่อใบ
การวิเคราะห์ transcriptional analysis ของทั้งจีโนม เผยให้เห็นว่าการตอบสนองอย่างรวดเร็วของต้นมะเขือเทศที่มีสภาวะขาดธาตุเหล็ก (Fe) นั้นได้รับอิทธิพลอย่างมากจากลักษณะตามธรรมชาติของตัวแปรต้นคีเลต (chelating agent) (Zamboni et al., 2016) อันที่จริง คีเลตธาตุเหล็ก-ไซเตรต (Fe-Citrate) และคีเลตธาตุเหล็ก-ไฟโตไซเดอโรฟอร์ (Fe-PS) ควบคุมการแสดงออกของยีน 728 คู่ และ 408 คู่ ตามลำดับ ผลวิเคราะห์นี้แสดงให้เห็นถึงการควบคุมกระบวนการนำส่งโมเลกุลลงตามลำต้น ซึ่งเกิดจากภาวะขาดธาตุเหล็ก (Fe) ในอีกด้านหนึ่งโครงสร้างเชิงซ้อนธาตุเหล็ก-ฮิวมิคที่ดึงแยกโมเลกุลน้ำได้ (Fe-WEHS) ไม่ได้เป็นปัจจัยสร้างการเปลี่ยนแปลงที่มีนัยยะสำคัญใน root transcriptome ในพืชที่มีภาวะขาดธาตุเหล็ก (Fe) ปรากฎการณ์นี้ชี้ให้เห็นว่ารากไม่ได้รับรู้ว่ามีการนำธาตุเหล็กมาเติมในส่วนที่พร่องไปในเซลล์ พฤติกรรมนี้จึงอธิบายการสะสมธาตุเหล็กที่เพิ่มขึ้นในพืชที่ได้รับปุ๋ยธาตุเหล็ก-ฮิวมิคที่ละลายน้ำได้
ผลของฮิวมิคต่อการเจริญเติบโตของรากและการทำงาน
การเติมฮิวมิคในพืชนำให้เกิดการเปลี่ยนแปลงในโครงสร้างรากและกำหนดกิจกรรมในชั้นผนังเซลล์ (Plant Membrane Activities) ที่เกี่ยวข้องกับการดูดซึมสารอาหาร เกี่ยวกับเส้นทางหลักและเส้นทางรองของการเผาผลาญพลังงาน ฮอร์โมน และปฏิกิริยาต่อสมดุลออกซิเจน (Reactive Oxygen Balance) (Varanini and Pinton, 2001; Nardi et al., 2002; Canellas and Olivares, 2014; Olaetxea et al., 2018; Figure 1) ผลกระทบเหล่านี้ แตกต่างกันไปตามแต่ที่มา ขนาดโมเลกุล และลักษณะทางเคมีของฮิวมิค ซึ่งแสดงให้เห็นบทบาทของเศษอินทรีย์วัตถุในการส่งเสริมการเจริญเติบโต และการต้านความเครียดในพืช
นักวิจัยหลาย ๆ ท่านสังเกตว่าพืชที่ได้รับฮิวมิคจากแหล่งที่ต่าง ๆ กัน ยังผลให้เกิดการแตกกิ่งรากและขนรากอย่างกว้างขวาง (Canellas et al., 2002; Nardi et al., 2002) พฤติกรรมนี้มีความเกี่ยวพันกับเส้นทางการส่งสัญญาณ (Signaling Path) ที่เกี่ยวข้องกับไฟโตฮอร์โมน โดยเฉพาะ ออกซิน ไนตริก ออกไซต์ Ca2+ และ ROS (Trevisan et al., 2010; Zandonadi et al., 2010; Mora et al., 2012; Ramos et al., 2015; García et al., 2016b,c) ทั้งนี้ การที่ยีนมีบทบาทควบคุมการดูดซึมขึ้นลำต้นของฮอร์โมนออกซิน (Trevisan et al., 2011) และการที่ฮิวมิคสามารถกำหนดรหัสพันธุกรรมสำหรับเอนไซม์ที่เกี่ยวข้องกับกระบวนการใช้ฮอร์โมน (hormone metabolisms) (Zanin et al., 2018) แสดงให้เห็นว่าฮิวมิคอาจมีอิทธิพลต่อความเสถียรของดุลยภาพฮอร์โมนพืชชนิดต่าง ๆ อย่างไรก็ตาม มีการสังเกตว่าการกระตุ้นการเจริญเติบโตของรากไม่มีความเกี่ยวข้องกับการเปลี่ยนแปลงของฮอร์โมน (Schmidt et al., 2007; Mora et al., 2012) สิ่งนี้สื่อให้เห็นว่า อาจมีสัญญาณอื่นเกี่ยวข้องกับการดัดแปลงโครงสร้างกายภาพที่เกิดจากสสารฮิวมิค
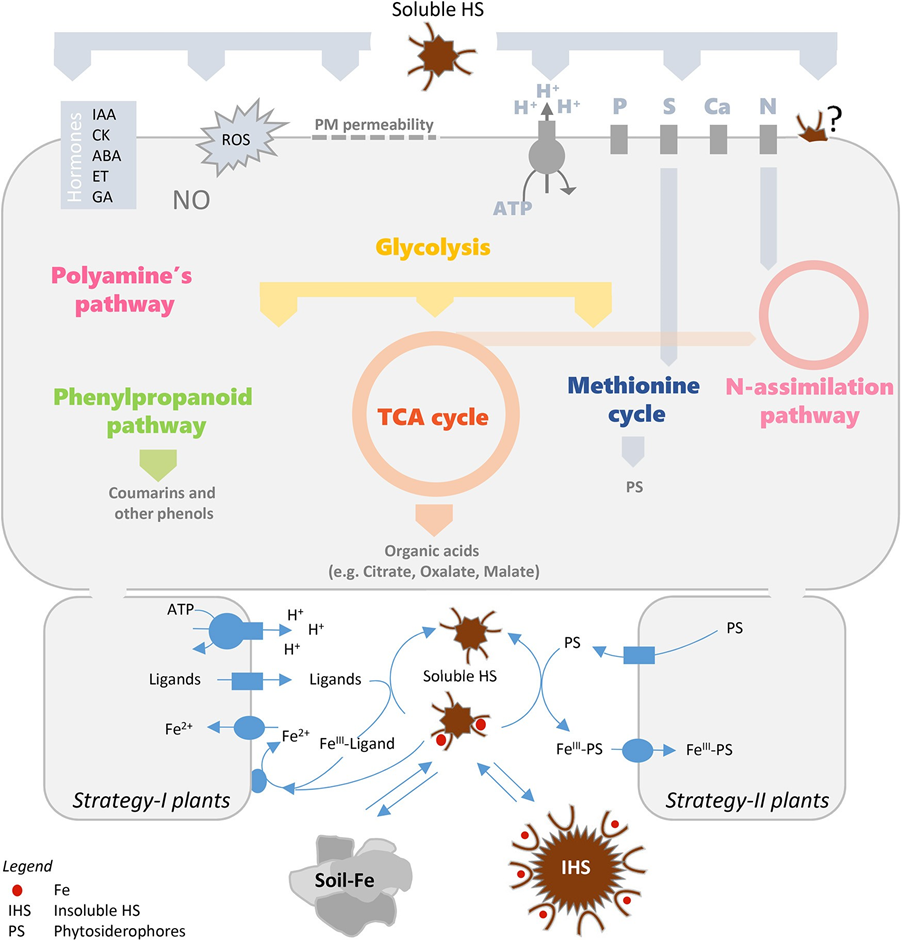
วงการวิชาการมองว่าปลายทางของกิจกรรมที่เกี่ยวข้องกับฮิวมิคคือ the root PM H+- ATPase (Zandonadi et al., 2016) มีการสังเกตหลักฐานการ activation of the PM proton pump ทั้งใน transcriptional and post-transcriptional level และที่เกี่ยวข้องกับโปรตอนที่ยื่นออกมา (extrusion) (Varanini et al., 1993; Canellas et al., 2002) และการดูดซึมไอออนจำพวกไนเตรต (Pinton et al., 1999; Quaggiotti et al., 2004; Tavares et al., 2017) ฟอสเฟต (phosphate) (Jindo et al., 2016) และซัลเฟต (Jannin et al., 2012) นอกจากการดูดซึมไอออนแล้ว ฮิวมิคยังเพิ่มการดูดซึมแร่ธาตุไนโตรเจนของพืช (Mora et al., 2010; Jannin et al., 2012; Vaccaro et al., 2015; Zanin et al., 2018) เพิ่มการเผาผลาญคาร์บอน (ไกลโคไลซิส และวัฏจักรเครบส์; Nardi et al., 2007; Trevisan et al., 2011) และการสังเคราะห์เมแทบอไลต์ (Metabolite) เช่น เฟนนิลโพรนอยด์ (phenylpropanoids) (Schiavon et al., 2010; Jannin et al., 2012; García et al., 2016c)
นอกจากการกระตุ้นการปล่อยโปรตอน ฮิวมิคยังมีผลต่อ rhizodeposition กรดฮิวมิคส่งเสริมการปล่อย anionic species close to บริเวณของ root acidification [apolar sugars from maize roots (Puglisi et al., 2009) กระทบต่อกลุ่มจุลินทรีย์ในดินในไรโซเสฟียร (Puglisi et al., 2013)] การเติบโตของรากที่เพิ่มขึ้นตามมาด้วยการปล่อยโมเลกุลน้ำหนักเบาจาก maize plants ที่ได้รับฮิวมิค (Canellas et al., 2019) ในอีกด้านหนึ่ง มีการรายงานว่ากรดอินทรีย์ อาทิ เช่น ที่ปล่อยมาจากรากพืช สามารถย่อยสลายโครงสร้างโมเลกุลขนาดใหญ่ กลายเป็นเศษฮิวมิคได้ (Piccolo et al., 2003) เศษฮิวมิคเหล่านี้จะส่งผลต่อรากอีกทีเช่นกัน ซึ่งในประเด็นนี้ เป็นที่น่าสนใจว่ามีการสะสมฮิวมิคที่ผิวรากและที่อะโพพลาสต์ (Apoplast) (García et al., 2012; Kulikova et al., 2014) นอกจากนี้ เศษฮิวมิคที่ได้จากดินไรโซเสฟียร (rhizospheric soil) ยังมีคุณลักษณะทางเคมีที่แยกออกจากดินอื่น ๆ (D’Orazio and Senesi, 2009)
ผลของฮิวมิคต่อการเจริญของราก (เส้นทางส่งสัญญาณ; signaling pathways) ต่อการดูดซึมไอออน (primary and secondary membrane transporters) ระบบเผาผลาญพลังงานหลัก (ไนโตรเจนและคาร์บอน), ระบบเผาผลาญพลังงานรอง (เฟนนิลโพรพานอยด์) และสารหลั่งจากรากพืช มีความสำคัญต่อการดูดซึมธาตุเหล็ก (Fe) และพัฒนาการตอบสนองของพืชต่อสภาวะการขาดธาตุเหล็ก (Fe) (Figure 1)
บทสรุปและมุมมอง
งานวิจัยจำนวนมากในทศวรรษที่ผ่านมาได้พิสูจน์ว่าศักยภาพของฮิวมิค เมื่อแยกจากแหล่งอินทรีย์วัตถุ มีผลต่อการเจริญเติบโตของพืช มีผลต่อสารอาหารพืช และการเผาผลาญพลังงานพืช
สสารเหล่านี้ ในดินธรรมชาติ มีความหลากหลายและการกระจายที่ซับซ้อน (heterogeneity and polydispersity) ทั้งสามารถตกตะกอนร่วมกับส่วนของแร่ธาตุต่าง ๆ (อาทิ เช่น เหล็กออกไซต์ – Fe-oxide และดินเหนียว) หรือในสารละลายที่มีผลต่อสารละลายอินทรีย์วัตถุปริมาณมาก
เนื่องจากมีน้ำหนักโมเลกุลที่ต่ำและเศษสสารที่ละลายน้ำได้มีผลต่อการทำงานของ Ion Transporters ที่ทำงานใน the PM of root cells ทำหน้าที่ในระดับ acting both at transcriptional and post-transcriptional level ผลการวิจัยนี้ได้มาจากการทดลองในสภาวะควบคุม เช่น การทดลองการแยกฮิวมิคและพืชที่ปลูกแบบไฮโดรโปนิคออกมา จึงเข้าใจได้ว่า ทำไมฮิวมิคจึงทำปฏิกิริยากับรากพืช และจุลินทรีย์ และเศษอนุภาคดินในไรโซเสฟียรได้ ดังนั้น การศึกษาลักษณะโครงสร้างและลักษณะทางเคมีของฮิวมิคในสารละลายดิน และในไรโซเสฟียรจึงมีความจำเป็น เพื่อที่จะถ่ายทอดองค์ความรู้ที่ได้จากการทดลองในสภาวะควบคุม ไปสู่การปฏิบัติงานในดินและไรโซเสฟียรทั่วไป ซึ่งจะชี้ให้เห็นถึงประโยชน์จากการวิจัยในการนฮิวมิคมาใช้ทำปุ๋ยสำหรับพืช และมีประโยชน์ต่อการส่งเสริมการเจริญเติบโตของพืช ต่องานวิจัยเกี่ยวกับความสัมพันธ์ระหว่างโครงสร้างทางเคมีและลักษณะของฮิวมิคที่ได้จากหลาย ๆ แหล่ง และมีโครงสร้างโมเลกุลที่มีความซับซ้อนแตกต่างกันไป ทั้งยังต่างกันในผลทางชีวภาพที่มีต่อพืช งานวิจัยเหล่านี้สามารถกระทำได้โดยใช้เทคนิคการวิเคราะห์ใหม่ ที่สามารถบันทึกลักษณะดั้งเดิมทั้งหมดขององค์ประกอบของฮิวมิคทุกชนิด ไม่ว่าจะเกิดขึ้นตามธรรมชาติ หรือเป็นมลภาวะที่เกิดจากฝีมือมนุษย์
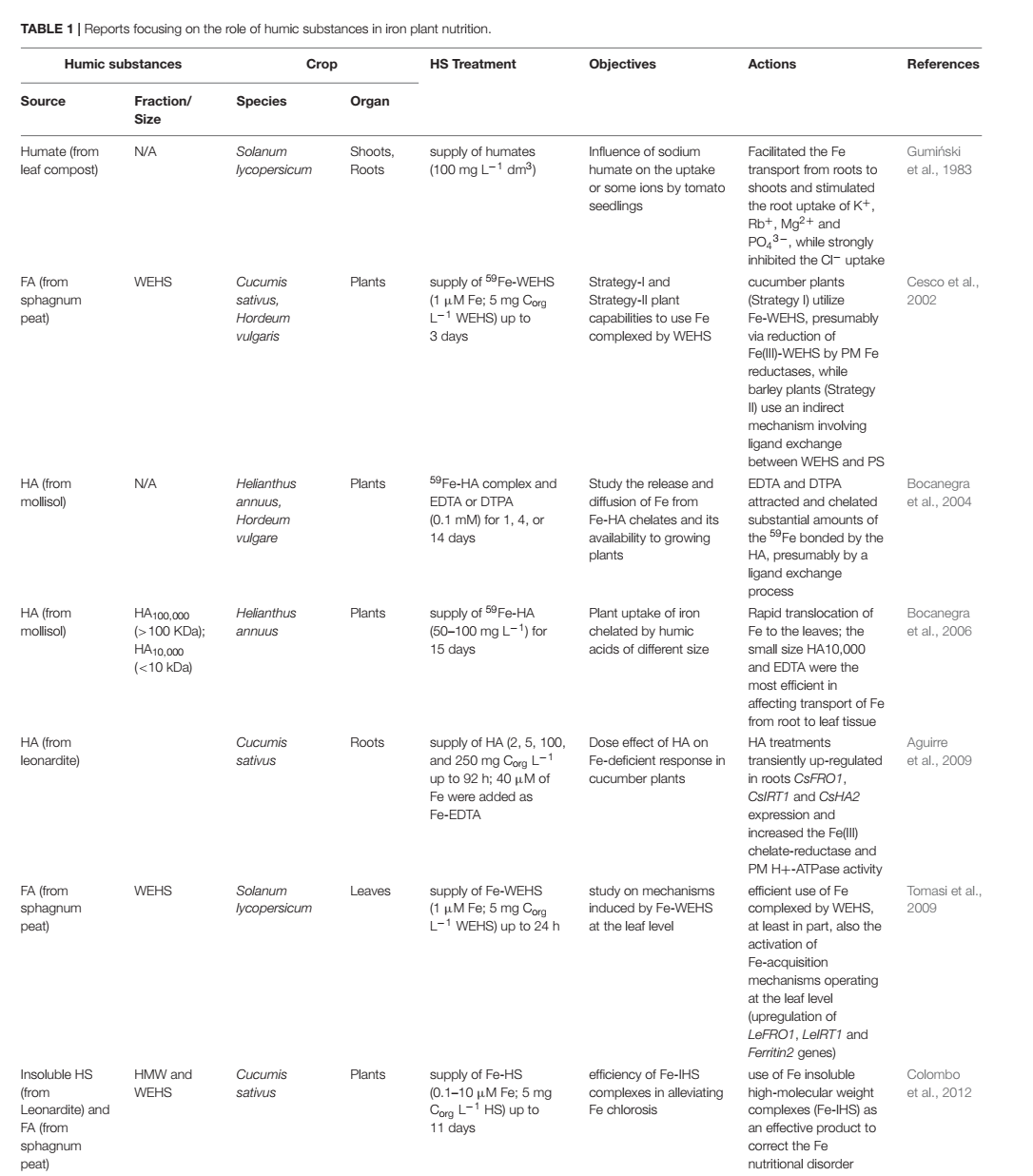
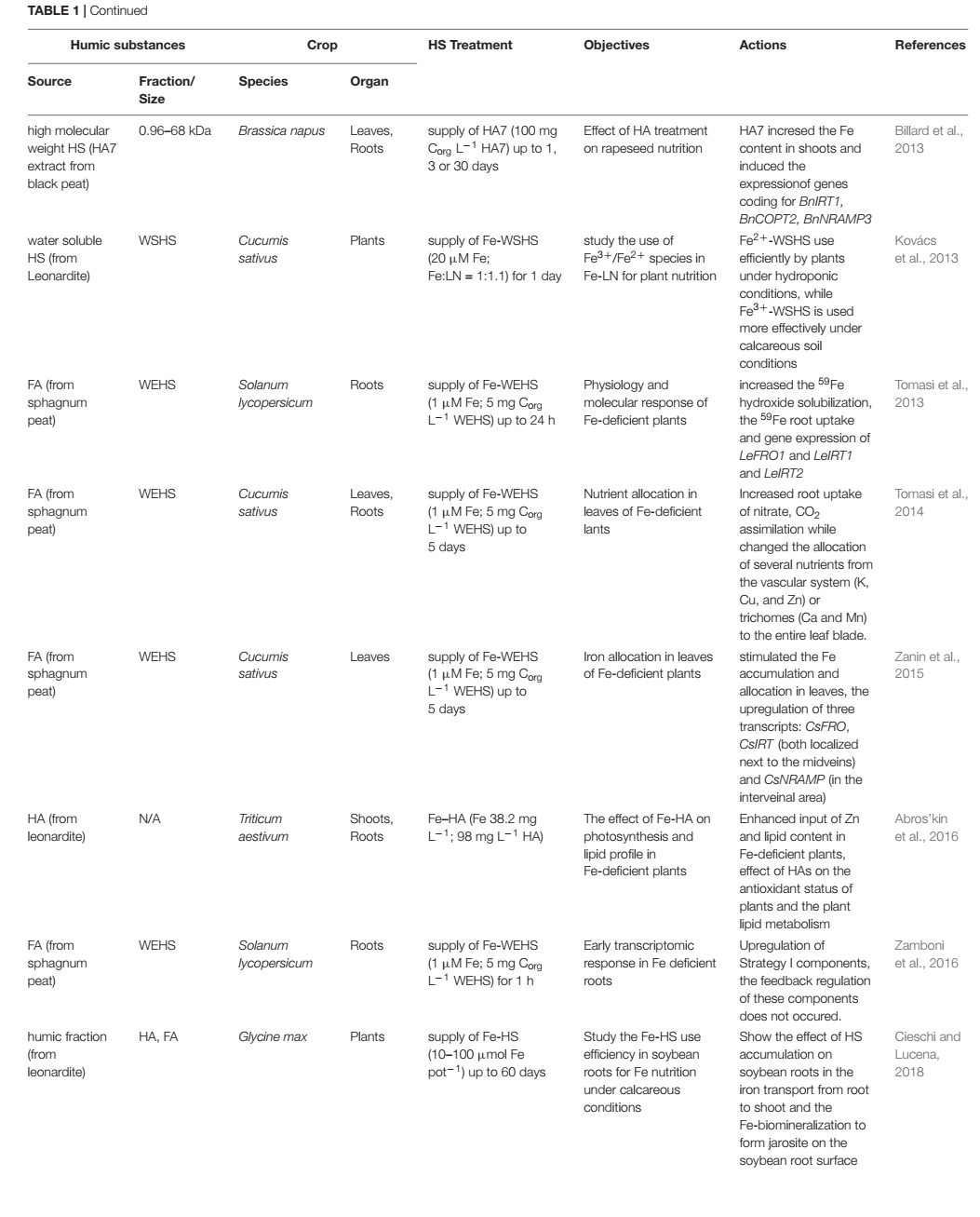
สารอาหารธาตุเหล็ก (Fe) มีความสัมพันธ์ที่เป็นประโยชน์อย่างยิ่งต่อบทบาททั้ง 2 ของสารฮิวมิค ทั้งในฐานะขององค์ประกอบของคีเลต และในฐานะของสารกระตุ้นชีวภาพ (Tale 1) เพราะศักยภาพของฮิวมิคในการก่อรูปโครงสร้างเชิงซ้อนกับธาตุเหล็ก (Fe) และผลต่อกระบวนการดูดซึมธาตุเหล็ก (Fe) แสดงถึงประโยชน์ของโครงสร้างเชิงซ้อนธาตุเหล็ก-ฮิวมิค (Fe-HS) ที่มีต่อสารอาหารธาตุเหล็กของพืช เมื่อเปรียบเทียบกับโครงสร้างเชิงซ้อนธาตุเหล็ก (Fe) อื่น ๆ ที่เกิดในไรโซเสฟียรตามธรรมชาติ
เป็นที่น่าสนใจว่าฮิวมิคนำให้เกิดการตอบสนองแบบ “nutrient acquisition response” แม้ว่าพืชจะได้รับสารอาหารเพียงพอแล้ว หรือได้ฟื้นฟูตนเองจากภาวะสารอาหารบกพร่องเรียบร้อยแล้ว ซึ่งการตอบสนองนี้กระทบต่อการทำงานและการควบคุมกระบวนการดูดซึมอาหารขึ้นไปใช้ รวมถึงโครงข่ายการส่งสัญญาณที่เป็นพื้นฐานของพฤติกรรมการดูดซึมนี้ นอกจากนี้ เป็นไปได้ว่าปฏิกิริยาระหว่างสารที่หลั่งจากรากและฮิวมิคอาจเป็นส่วนหนึ่งของการสื่อสารระหว่างพืชและดิน ปฏิกิริยาไขว้กันเหล่านี้นำให้เกิดการปรับตัวของพืชต่อสิ่งแวดล้อมหนึ่ง ๆ
อีกประเด็นหนึ่งที่น่าสนใจในงานวิจัยนี้ที่เกี่ยวข้องกับพฤติกรรมของฮิวมิค คือประโยชน์จากการนำไปพัฒนาปุ๋ยที่เป็นมิตรต่อสิ่งแวดล้อม ซึ่งมีความสำคัญอย่างยิ่งในเศรษฐกิจหมุนเวียน แม้ว่าโครงสร้างทางเคมีของฮิวมิคจะยังไม่เป็นที่กระจ่างชัด และการนำผลทดลองไปปรับใช้ในดินจริง ๆ ยังเป็นที่กังขา แต่ก็ชัดเจนว่าฮิวมิคที่แยกจากอินทรีย์วัตถุอื่น ๆ เมื่อเติมเข้าไปในสารละลายอาหารพืช หรือเติมเข้าไปในดิน จะเพิ่มคุณค่าให้กับสารอาหารนั้น ๆ และมีประโยชน์อย่างยิ่งกับการดูดซึมไนโตรเจน และธาตุเหล็ก สิ่งนี้สื่อให้เห็นว่าเศษฮิวมิคที่มีคุณสมบัติทางเคมีและชีววิทยาต่างกัน สามารถใช้ออกแบบปุ๋ยประสิทธิภาพสูง โดยใช้ฮิวมิคเป็นพื้นฐานได้ สิ่งนี้จะมีความสำคัญในการทำงานกสิกรรมแม่นยำ ที่มุ่งเป้าไปยังการจำกัดการใช้ทรัพยากรณ์จากภายนอกระบบนิเวศน์ธรรมชาติ และเพิ่มการใช้ทรัพยากรณ์ธรรมชาติ
แปลจาก
Humic Substances Contribute to Plant Iron Nutrition Acting as Chelators and Biostimulants
คำสำคัญ
Abbreviations: ABA, abscisic acid; CK, cytokinins; DOM, dissolved organic matter; ET, ethylene; FA, fluvic acids; FRO, ferric chelate reductase; GA, gibberellic acid; HA, humic acids; HS, humic substances; IAA, indole-3-acetic acids; IHS, insoluble HS; IRT, iron transporter; NO, nitric oxide; NRAMP, natural resistance-associated macrophage proteins; PS, phytosiderophores; ROS, reactive oxygen species; TCA, tricarboxylic acid; WEHS, water extractable humic substances.
